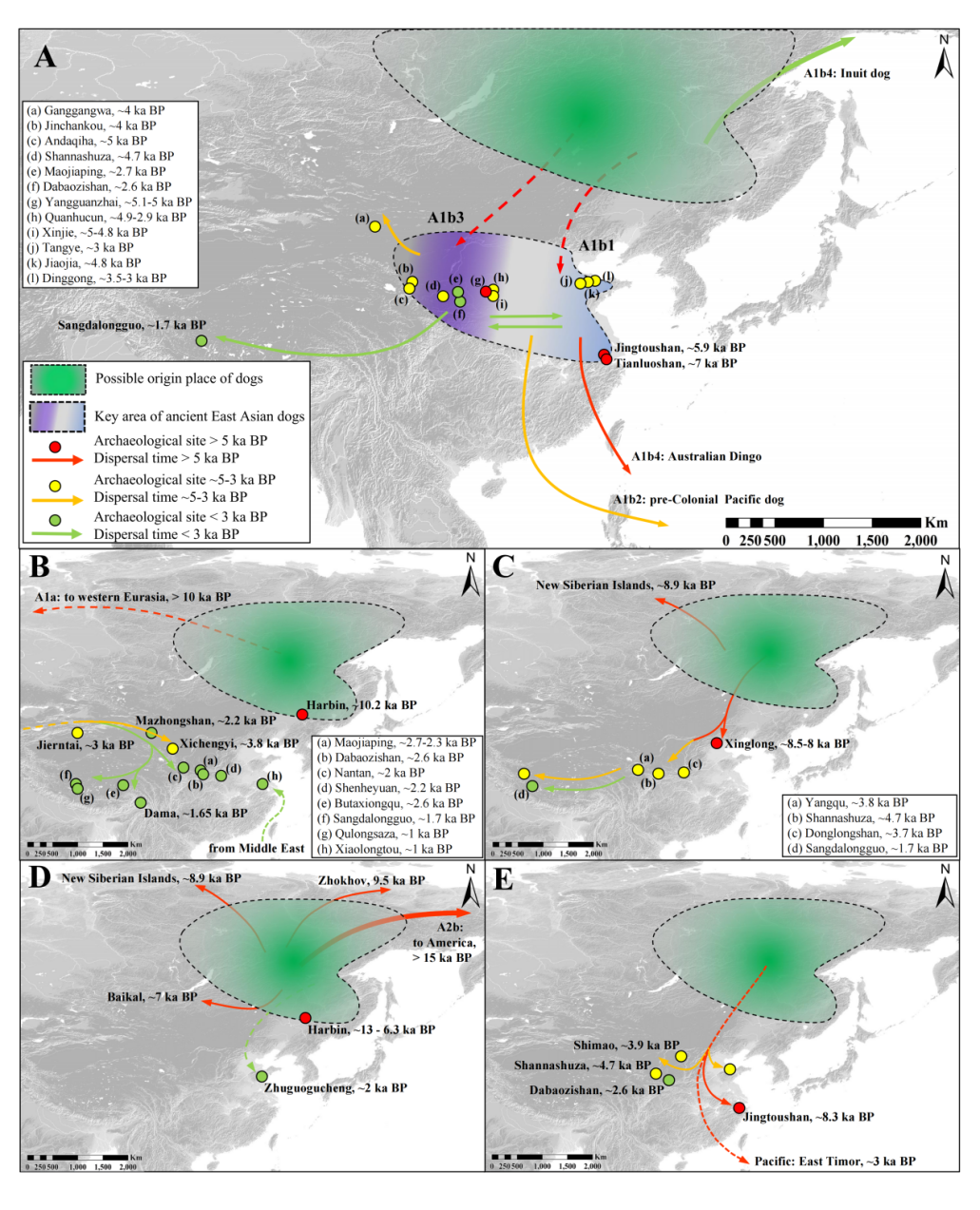
5 f% v8 j) ` B# O4 x5 B* {另一篇原文! k; [, h6 c/ G5 H; J https://mp.weixin.qq.com/s/1M4mUywoePeYRciKTScrDQ 9 d% i( O- G) p- `4 |2 J$ m6 L9 \* {夏依热·肖开提 | 古DNA提取技术的优化及其应用简论/ ?: J' }% ?; S7 V
夏依热·肖开提 中国社科院考古所中国考古网 2024-04-17 16:53 北京 V4 N1 q; k$ n* @- @摘要:随着古DNA技术的不断突破与进步,其在动物考古学领域中的应用也日益广泛。通过分析考古遗址出土的动物遗存中提取的古DNA,结合传统考古及动物考古学研究的成果,为我们揭示家养动物的起源、驯化和扩散以及人类与动物共同演进的历程提供了重要的线索。然而,古DNA研究受其样本的年代及遗址所处环境的影响较大。特别是南方地区的酸性土壤环境会严重侵蚀和破坏骨骼结构,这对DNA的保存和提取极为不利。因此,在进行古DNA研究时,我们必须充分考虑环境因素对DNA保存状况的影响,并采用相应的技术手段,最大限度地优化古DNA的提取过程,以确保研究的准确性与可靠性。 % T, a7 v: y3 F2 H' Z/ u( R; R 4 ~4 i* f8 F" N) A$ X% a. }: U) I! ?: J" s
6 _7 j1 k' B& D v
古DNA研究已成为科技考古领域中一门蓬勃发展的学科。通过分析古生物遗存中的DNA,我们能够揭示物种的进化历程、人类迁徙及文化交流等方面的信息。自20世纪80年代以来,学者们一直试图从古生物的遗存中提取DNA,但由于古DNA的高度降解和污染问题,早期的研究进展缓慢。直至20世纪90年代中后期,随着分子生物学技术的发展,特别是聚合酶链式反应(Polymerase Chain Reaction,PCR)的应用,古DNA研究领域取得了突破性进展,并逐渐成为一门内容丰富的学科,涉及领域也逐渐广泛,如人类的起源、动植物的驯化和扩散传播等。进入21世纪后,高通量测序技术(Next generation sequencing,NGS)的出现使得学者们能够更准确地测定和分析古生物的DNA序列,从而揭示出更多关于物种进化和人类历史的信息。在这一波技术革新的推动下,古DNA研究迈入了一个全新的时代。2022年,瑞典古生物学家斯万特·帕博(Svante Pääbo)因其在已灭绝的人类基因组和人类进化方面的杰出贡献而被授予诺贝尔生理学或医学奖,更是将古DNA研究推向了生命科学领域的前沿。8 m* I) A8 M, | ?. y, N* i
家养动物的起源与演化一直是考古学和遗传学领域的研究热点。古DNA在动物考古学中的广泛应用,为我们提供了全新的视角和研究手段,使我们更深入地理解动物的驯化与扩散以及古代社会人类与动物之间的互动关系。然而,需要注意的是,古DNA研究仍面临许多挑战和限制,如样本的获取和保存、DNA的降解和污染等问题。尤其是南方地区由于其埋藏环境等因素导致出土动物骨骼样本的保存状况普遍不佳,保存完好的动物遗存标本数量稀少。这无疑给南方地区的动物古DNA研究带来了严峻的挑战。研究者们需要面对如何从稀有、保存质量差且掺杂大量污染影响的样本中,有效提取出足量、高质量的古DNA等问题。这就涉及到考古遗址出土动物遗存的古DNA提取技术的完善以及其研究思路的改进问题。本文就此略作简论,以期抛砖引玉可以促进学界就相关问题展开更多的讨论,推进考古遗址尤其是南方地区遗址出土动物遗存的古DNA样本提取技术与研究的改进。/ I* N% E2 V8 v& e
一、古DNA提取技术的优化) w0 f& j/ F9 u! E# b3 o! F1 Z
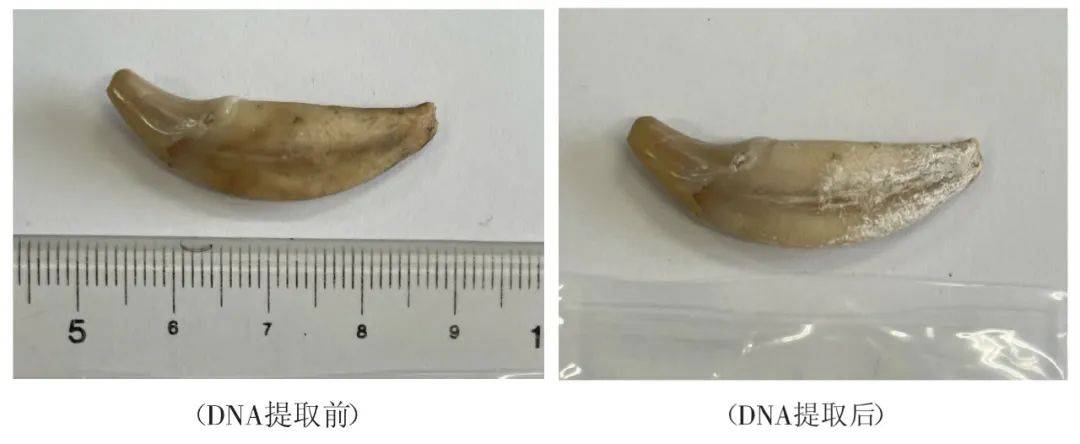
古DNA研究面临的主要挑战在于,由于样本的年代久远以及埋藏环境等多种因素作用,古DNA会发生不同程度的降解和污染,因此会导致提取到的古DNA存在片段缺失、碎片化以及参杂大量污染,即内源DNA含量低。这些挑战对古DNA的提取和后续分析都提出了更高的要求,需要研究者采取更为精细和严谨的实验方法和技术手段来应对。其中环境因素在古DNA的保存过程中起着决定性的作用。古DNA的降解速率与其保存环境如温度、湿度和pH值等因素密切相关。北方地区大多数遗址低温、干燥、弱碱性及微生物活动较少的封闭环境有效地降低了对DNA的破坏作用,从而为古DNA的长期保存提供了相对有利的条件。而南方地区多数遗址出土动物骨骼样本保存情况普遍不好,主要是因为南方地区高温和潮湿的气候特征利于微生物的生长繁殖,从而加速古DNA降解。此外,南方地区的酸性土壤不利于古DNA的保存,其酸性土壤会破坏DNA的结构,导致DNA损伤。综上所述,南方地区高温、潮湿及酸性土壤等气候环境因素相互作用,加速了古DNA的降解和损伤过程,给古DNA的提取和研究带来了极大的挑战和困难。因此,在南方地区进行古DNA研究时,如何获取高质量的古DNA样本,如何从大量杂质中提取出微量的古DNA,并且确保提取到的DNA信息的完整性和可靠性是古DNA研究中的关键性问题。 ' h4 Y7 q. c& o- G2 }目前古DNA提取多数是从颞骨或者牙齿进行,这是因为颞骨是头骨中最致密、最坚硬的部位之一,能够很好地保护内部的DNA不受外界环境的破坏[1]。而牙齿的特殊结构使得其内部的DNA受到较好的保护。研究发现,牙骨质中的线粒体DNA(Mitochondrial DNA, mtDNA)含量比牙本质高五倍,这使得富含牙骨质的根尖成为古DNA研究中极为理想的取样部位之一[2]。随着技术的不断进步,学者们也在不断改进和优化古DNA提取方法。笔者在读博士课程期间致力于优化古DNA提取方法,为了最大程度地减少对珍贵样本的损害(如骨骼或牙齿结构等),笔者积极推动采用微量牙骨粉(图一)及微损伤法(Minimum distractive method)(图二)来提取古DNA。相较于传统的古DNA提取方式,即依赖于钻孔、切割以及对(部分)骨骼和牙齿进行粉末处理。微损伤法[3]是将牙齿的部分牙根直接浸泡在裂解缓冲液中,并在不破坏牙齿结构的前提下,从牙根的牙骨质中分离出古DNA的提取方法(图二)。此方法与传统的磨粉取样相比,也可以提取等量古代DNA。这种方法不需要对样本进行破坏性钻孔或研磨,并且在提取后,其牙齿仍然可以用于多数形态学研究。如图一展示了日本最古老的家犬之一(日本爱媛县上黑岩遗址出土,绳文时代,7400~7200 cal.BP)取样前后对比。鉴于样本的珍稀性,我们采用了在尽量不影响牙齿完整结构的前提下,磨取微量牙骨粉(~35mg)来提取古DNA。随后,我们将提取到的DNA通过线粒体捕获技术进行了进一步的分析和处理。图二展示了日本弥生时代出土家犬样本DNA提取前后对比。这是我们首次尝试通过微损伤法来提取古DNA。经过后续纯化和测序检测,显示此方法能够成功提取到较高含量的内源DNA,为古DNA研究提供了新的有效手段。 ( w$ J$ N' m9 P! e5 Z+ R 7 w) n' g% K$ I5 D; |6 O* m" t9 H! M) T d& P
图一 日本绳文时代上黑岩遗址出土家犬微量牙骨粉取样前后对比 ; y6 z3 `" t+ ?7 X0 E 3 ^% _7 C% Y& H. r# A5 i5 x* `1 j 7 ?8 z: m+ A' z( t$ H5 z图二 日本弥生时代出土家犬微损伤法 DNA 提取前后对比 - X& Z% k1 R# l. ? & d3 h- ~' x9 u% n4 N. o# m鉴于日本列岛与我国南方地区埋藏环境的相似性,笔者认为,在国内处理那些难以从骨骼中提取或足量提取古DNA的样本,我们也应积极采用微量牙骨粉或微损伤法即从牙根的牙骨质中提取古DNA,在降低对样本的破坏度的同时,保证提取成功率。对于骨骼保存不好或数量有限的考古遗址特别是对于南方地区重要遗址中出土的极为重要的动物样本,我们应综合埋藏环境,依据样本实际情况制定合适的提取、捕获和测序方案。从测序技术层面而言,不断进步的测序技术及基因组数据分析方法进一步提高了古DNA研究的准确性和分辨率。例如,古DNA捕获技术,通过设计特定的DNA探针,能够高效地从大量的污染DNA中捕获出目标DNA,从而大幅度提高了古DNA的测序效率,并有效地从一些难以处理的样本中得到有用的数据用于分析[4]。 ' n/ g8 P1 J& D2 U二、动物古DNA研究的探讨 ! I) v( i5 Y/ j9 J利用古DNA技术从古代动物尤其是家养动物的遗存中提取遗传信息(包括线粒体、多态性位点及全基因组数据等),可以为我们揭示其遗传背景、种群结构、亲缘关系及家养动物从驯化中心向其他地区扩散的情况,以及与当地野生种群杂交的情况。这将为我们理解家养动物的驯化过程和遗传多样性提供重要线索。国际上,对于家养动物如狗、猪、马等的起源地和时间已有不少研究。这些研究借助古DNA分析技术深入揭示了家养动物的遗传演化历程和与人类文明的互动关系。在国内,随着考古发掘工作的不断深入和古DNA技术的日益成熟,越来越多的学者开始关注中国家养动物的起源问题,并取得了一系列重要成果。例如中国社会科学院、中国科学院、吉林大学、复旦大学等研究机构对中国主要家畜猪、马、黄牛等进行了古DNA分析,为研究中国古代家养动物的起源与扩散提供了宝贵的依据[5][6][7][8][9]。笔者认为古DNA研究在动物考古学中的应用具体主要体现在以下两个方面:(1)古DNA研究可以帮助我们了解家养动物的驯化和扩散过程,进而揭示人类与动物之间的互动关系。同时,通过比较家养动物DNA与野生物种的差异,以及不同地区家养动物DNA的遗传差异,可以推断动物的迁徙和传播路线;(2)古DNA研究还可以揭示家养动物种群的历史变化,包括种群大小、遗传结构及多样性以及一些表型特征等,这对于我们了解动物的生物学特性及生态适应性、人类对动物的开发利用等活动以及人与动物的关系等具有重要意义。: _1 D. C/ f. u& K
以家犬的古DNA研究为例,笔者的博士论文主要围绕东亚地区(主要是日本列岛)不同时期出土的家犬为研究对象,从古DNA视角出发(线粒体和全基因组),通过结合考古材料背景、形态学研究来进一步详细讨论了东亚地区家犬的驯化及与人类的关系。一直以来,家犬的起源都是个比较有争议的话题。先前的研究中,有学者认为家犬的起源在东亚,有的则认为是在欧洲,且有研究者认为家犬在东亚及欧洲地区是被独立驯化的[10][11][12][13][14]。因此,家犬的具体起源时间、地点和驯化方式等仍存在较大的分歧。目前,关于家犬的古DNA研究也都集中在来自欧洲、近东和西伯利亚的样本上。在国内,古代家犬的研究工作也取得了一定进展。中国科学院和吉林大学的团队已分别对26只和29只古代家犬的线粒体基因组进行了探究,揭示了中国古代家犬的母系遗传特性[15][16]。然而,我们还需要大量东亚的古代犬和狼的基因组信息和考古学证据来进一步探讨其起源地。& _/ H) K1 N$ y1 C5 t7 v
笔者在读期间,通过分析12只日本古代犬(包括5只绳文时代,7400~4000年前;7只奈良时代,7~8世纪)的基因组数据,首次揭示了日本古代家犬的遗传特性[17]。基于线粒体基因组的研究结果,日本绳文犬(来自三个不同时期、不同地点出土的样本)在进化树上形成了一个单系群。该单系群与由中国现生南方犬以及泰国犬所构成的亚支系为姐妹支系。此外,绳文犬亚支系位于以浙江田螺山出土家犬为中心的网络结构图上基部附近[18]。这些发现引发了关于绳文犬可能起源的讨论。尽管存在绳文犬可能从中国南方(绳文时代早期)引入的假设,但考虑到田螺山个体的年代(约7000年前)晚于绳文犬的推测传入日本列岛的时间(11500~9300年前),目前尚无法断定绳文犬的确切起源地。为了更深入地探讨这一问题,我们需要获取更多来自中国,尤其是华南地区的古代犬类样本。此外,近期发表的关于日本古代狼的全基因组研究也为家犬起源的讨论提供了新的视角。该研究指出,家犬很可能起源于东亚地区,并特别强调了尤其是中国古代家犬和古代狼在解决家犬起源问题上具有重要性[19]。这些古DNA分析为深入探讨家犬的起源和进化提供了新的线索和研究视角。然而,要确切揭示家犬的起源问题仍需更多的基因组数据和考古学证据的支持。因此,笔者认为,通过优化古DNA提取技术,扩大东亚地区的古代犬和狼的基因组研究样本范围,深度测序及更为深入的分析,我们能够更全面、更准确地探究家犬的起源、进化和扩散等过程。 + g5 p$ W9 o5 @- ~1 U- N {. H除此之外,笔者还通过从全基因组数据及其提取到的多态性位点信息分析了绳文犬与奈良时代日本古代家犬的一些遗传特性,包括性别,体型大小,耳朵形态,淀粉消化能力等。随着研究的不断深入,其对于进一步理解家犬的遗传特性以及其在不同历史时期的演化过程具有极其重要的学术意义。+ E. n3 X5 {+ E# `6 {/ i
家犬作为人类最早驯化的动物之一,其古DNA研究为我们揭示家犬的起源、理解家犬与人类之间密切的关系提供了宝贵线索。鉴于此,我们应该将古DNA研究的范围扩展到更多的动物种类中,例如牛、羊、鸡等这些在人类农业文明的发展过程中扮演了重要角色的家养动物。同时,野生物种乃至已灭绝动物的古DNA研究同样具有重要意义。它们可以帮助我们重建史前生态环境,理解物种间的相互关系及其对环境变化的适应策略等。! K$ R6 w# q$ c, a$ `$ n; k$ v
三、结 语8 l2 Y$ z; h/ W# N7 i2 M$ h3 ]/ j
随着技术的不断进步,笔者坚信古DNA研究在未来将会迎来更加广阔的发展空间。首先,从技术层面来看,古DNA提取方法、捕获和测序技术以及分析方法的持续进步,为动物古DNA研究带来了革命性的变革。这些技术的不断创新和完善,使得我们能够更加高效、准确地提取和分析动物遗存中的古DNA信息。与此同时,针对考古遗址出土动物遗存的埋藏环境、骨骼保存情况等复杂因素,我们也应该不断探索和优化古DNA样本提取技术,以最大程度地克服环境等因素的干扰,提高古DNA研究的准确性和分辨率;其次,跨学科多领域合作将会为古DNA技术在动物考古学领域的研究提供更为全面的信息。例如,同位素分析可以检测家养动物骨骼或牙齿中的稳定同位素(如碳、氮、氧、锶等)比值,这些比值反映了动物的食性和生活环境。通过对比不同地区和时间点的同位素数据,可以推断出家养动物的迁徙模式和生态适应策略。而经典动物考古学能为我们提供家养动物的考古学背景,包括动物种群的物种组成、体型变化、种群结构、埋葬形式等。综上所述,将古DNA研究与同位素分析、动物考古学相结合,可以为我们提供一个多维度的视角,去探讨家养动物的起源、驯化及迁徙历程,也有助于更深入地理解家养动物在古代社会中的应用功能及其意义以及它们与人类之间的互动联系。这种跨学科的研究方法将为家养动物考古学研究带来全新的突破和发现。 ; H0 \# e6 x( c% ?+ N古DNA研究作为科技考古领域的一门新兴学科,已经为我们揭示了关于动物驯化和扩散、物种遗传特征等诸多方面的信息。在未来的发展中,随着技术的不断进步,我们有理由相信古DNA研究将在动物考古学中继续发挥重要作用,为我们深入探索理解动物驯化历程、人类与动物的关系、古代生态以及社会环境等提供宝贵的信息。 2 L# j1 H2 {8 m c附记:本文为国家重点研发计划资助“公元前1500年至公元前1000年中华文明早期发展关键阶段核心聚落综合研究·商代都邑的环境、生业与人群”(课题编号2022YFF0903605)研究成果、“河南省四个分时期专题历史文化研究课题”中“夏文化的生业、资源与经济技术”课题、中国社会科学院创新工程项目“古代动物的驯养与开发利用——动物考古的多学科研究”(项目编号2024KGYJ013)的阶段性成果。感谢日本综合研究大学院大学寺井洋平副教授对本文的学术支持。4 X* O6 z7 I! P3 u1 B9 [. q8 @
/ F0 X- A c T; r! h0 K6 S: y
( T( \% d, m9 V Z+ `- D& Y向上滑动阅读注释 ; @ [1 @; x2 K- y8 L8 L4 z: X' \) U7 ^, P! a+ ~: ^. W
[1]Ron P, Daniel F, Kendra S, et al. 2015. Optimal Ancient DNA Yields from the Inner Ear Part of the Human Petrous Bone. PLoS ONE, 10(6), e0129102. , ]0 H2 j9 ?6 Z2 u
[2]Adler C J, Haak W, Donlon D, et al. 2011. Survival and recovery of DNA from ancient teeth and bones. Journal of Archaeological Science, 38, 956-964. ) I6 v3 W7 E7 E+ ?- G! ^8 j+ m[3]Harney A, Cheronet O, Fernandes D M, et al. 2021. A Minimally Destructive Protocol for DNA Extraction from Ancient Teeth. Genome Research, 31, 472-483. ) b: M# T8 j E+ }. i# H( F
[4]Bahcall,O. Capturing ancient DNA. 2013. Nature Genetics, 45, 1417. 9 J8 u$ C& O0 t: G[5]崔银秋、张雪梅、汤卓炜等:《家猪起源与古代DNA 研究》,《边疆考古研究》2010年第1期。! B9 X1 ^- `# r% b% ]% E# S
[6]赵欣、李志鹏、东晓玲等:《河南安阳殷墟孝民屯遗址出土家养黄牛的DNA研究》,《第四纪研究》2020年第2 期。 ' W8 J& m5 m; F[7]文少卿、俞雪儿、田亚岐等:《古基因组学在古代家马研究中的应用》,《第四纪研究》2020年第2期。 $ @' M6 e& E9 X
[8]蔡大伟、孙洋:《中国家养动物起源的古DNA研究进展》,《边疆考古研究》2012年第2期。 1 {* F4 k5 y* N; S: M9 R
[9]Zhang M, Sun G, Ren L, et al. 2020. Ancient DNA evidence from China reveals the expansion of Pacific dogs. Molecular biology and evolution, 37, 1462-1469. $ S4 E! S- t7 j9 w' q
[10]Savolainen P, Zhang YP, Luo J, et al. 2002. Genetic evidence for an East Asian origin of domestic dogs. Science, 298, 1610-1613. ' D. n, ^, o' J, O
[11]Pang JF, Kluetsch C, Zou XJ, et al. 2009. mtDNA data indicate a single origin for dogs south of Yangtze River, less than 16,300 years ago, from numerous wolves. Molecular biology and evolution, 26, 2849-2864. 3 R: Z f5 i' d6 D; k' Y! `
[12]Thalmann O, Shapiro B, Cui P, et al. 2013. Complete mitochondrial genomes of ancient canids suggest a European origin of domestic dogs. Science, 342, 871-874. - `$ Y1 |- w9 O. Z! k[13]Frantz LA, Mullin VE, Pionnier-Capitan M, et al. 2016. Genomic and archaeological evidence suggest a dual origin of domestic dogs. Science, 352, 1228-1231.. Y+ A4 T7 {! E4 c
[14]Bergstroöm A, Stanton DW, Taron UH, et al. 2022. Grey wolf genomic history reveals a dual ancestry of dogs. Nature, 607, 313-320. # P1 |0 ], \% p3 d9 {[15]同[9] . G Z( d' \; y4 n J5 R1 B[16]Wang XC, Wei WQ, Zhang NF, et al. 2024. Ancient dog DNA reveals the human live-lihodd mode trasitions during the late Neolithic in Northeastern China. Journal of Archaeological Science: Reports, 53, https://doi.org/10.1016/j. jasrep.2023.104349. - g+ j1 L# n, M( R" z3 K3 L[17]Xiaokaiti X, Sato T, Kasai K, et al. 2023. The history of ancient Japanese dogs revealed by mitogenomes. Anthropological Science, 132, 1-11. $ P3 l; k+ P1 b7 _
[18]同[17 ] + S1 c( A1 }# ~% F& {. T[19]Gojobori J, Arakawa N, Xiaokaiti X, et al. 2024. The Japanese wolf is most closely related to modern dogs and its ancestral genome has been widely inherited by dogs throughout East Eurasia. Nature Communications, 15, https://doi.org/ 10.1038/s41467-024-46124-y. # i# _! J3 Q, P( \/ {( n$ P5 r* A( H Y+ P2 J
作者:夏依热·肖开提(中国社会科学院考古研究所)* K% j; M, g; z; z
 ! A8 H3 Q0 _! v! [& r* t& D! b, Z0 ?
! A8 H3 Q0 _! v! [& r* t& D! b, Z0 ?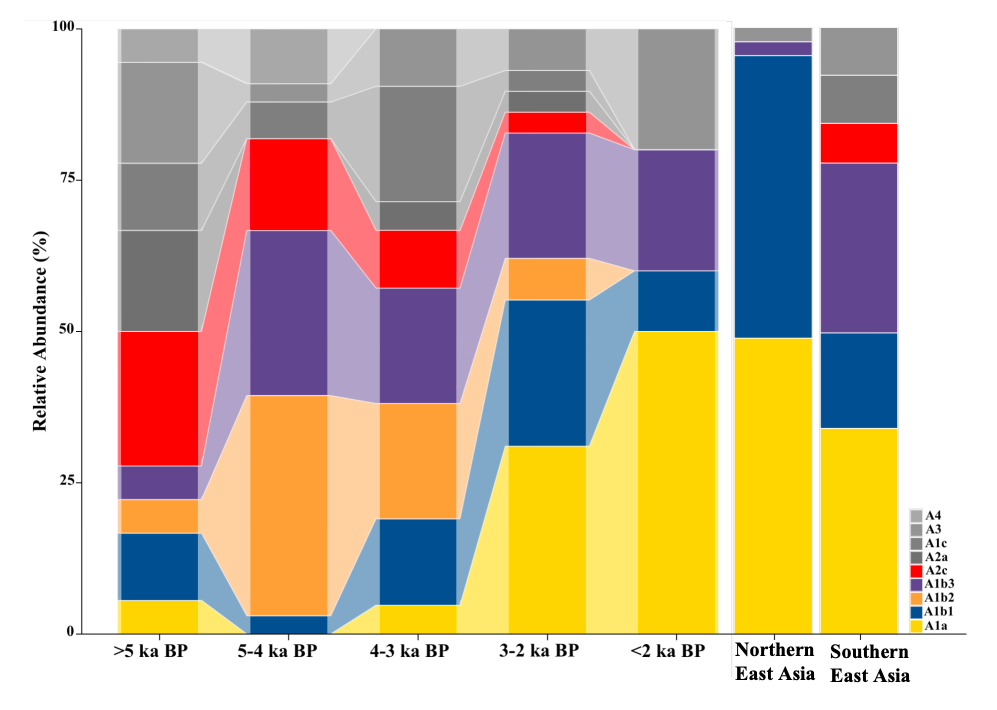 + Q% I* a9 A. A+ R
+ Q% I* a9 A. A+ R